The Dentinger lab supports a wide array of research including graduates and undergraduates investigating fungal biodiversity and evolution. Our research has been supported by NSF, National Geographic, and through generous donations.
BRYN DENTINGER
Curator of Mycology and Associate Professor, Natural History Museum of Utah and the School of Biological Sciences, University of Utah
Bryn hails from Minnesota and attended the University of Minnesota for his PhD, where he studied the molecular systematics of clavarioid and porcini mushrooms. He has carried out fieldwork all over the world, including exciting collecting trips to Vietnam, Brazil, and Cameroon. Outside of work, Bryn enjoys cooking (preferably with mushrooms) and brewing beer, showing that even his hobbies often involve a mycological component.
Colin is a Bay Area California native whose research focuses on unravelling the ethnomycological mystery of psychoactive bolete mushrooms. With a lifelong interest and undergraduate degree in astronomy before changing his studies to the ethnobiology of psychoactive substances, Colin has always felt attracted to explore the mind-bending wonders of nature. In his free time, he enjoys engaging in just about any outdoor activity with friends, most of all foraging for mushrooms.
Kendra is an Oregonian whose research interests center around symbioses, coevolution, and genomics. Her favorite mushroom is the lobster mushroom (not really a mushroom, but a parasitic mold attacking a mushroom host!). She only has one fungal tattoo, for now–a Hypogymnia lichen on her forearm. Kendra is currently a postdoctoral researcher with the Science Research Initiative at the University of Utah.
Keaton is an Idahoan whose research focuses around population genomics, speciation genomics, and ectomycorrhizal ecology. His favorite mushroom is the noble porcini. Keaton and his mustache enjoy having outdoor adventures with his fiancée and their Boston terrier, Evie. Keatong is currently a postdoctoral researcher with Rytas Vilgalys at Duke University.
Alex is a Northern California native whose research projects include investigating genetic diversity in large fungal individuals and metabarcoding of vertebrate scat samples to characterize diet, microbiome, and fungal distributions. He is a devoted owner to his merle longhair dachshund Bones. Alex is currently a postdoctoral researcher with David Hibbett at Clark University.
Bryce is from Utah and has worked on projects involving metabarcoding fungal propagules carried on bird feathers and mining transcriptomic datasets for hidden mycoviruses.
My lab studies fungal diversity using collections-based research, integrating fieldwork and molecular data to discover and understand the awesome and often weird world of Fungi. We routinely employ DNA barcoding to identify fungi collected from all over the world. More recently, genome sequencing has become standard practice for phylogenetic inference, metagenomic discovery in complex samples, and comparative approaches to elucidate natural product biosynthesis and genome evolution.
Projects
Diversity and Evolution of Ectomycorrhizal Fungi in the Guineo-Congolian Rainforest
Funded by NSF (DEB#1556412), 2016-2019; Terry Henkel (PI), Cathie Aime (Co-PI), Bryn Dentinger (Co-PI)
Sorting collections from a plot survey in Gilbertiodendron dewevrei-dominated forest in the Dja Reserve, Cameroon
Some fungi live in close symbiotic association with plants. These ectomycorrhizal fungi provide resources to the plant host, and in return, the plant provides nutrition to the fungi. The interactions of fungi and their hosts provide crucial ecosystem functions including the recycling of nutrients. While the critical roles that fungi play in the environment are well established, their importance in tropical systems is not well understood, and this is especially true for tropical Africa. This project in the Guineo-Congolian rainforest will provide the first thorough documentation of these fungi and plant relationships of tropical Africa using modern methods. The huge forest covering the Congo River basin is one of the last great tropical rainforests of the world. Current threats to the region include logging, mining, and climate change. This project will fill a major gap in our knowledge of the important symbiotic fungi of this ecosystem through expeditionary fieldwork in the undisturbed rainforests of Cameroon. This project is potentially transformative in that if the fungi associated with closely related host plants of Africa and South America are shown to have a common ancient origin, such results would fundamentally impact understanding of the evolution, global biogeography, and local adaptation of ectomycorrhizal symbioses. Additionally, application of new DNA sequencing methods will develop entirely new, phylogenetically informative data that will transform the field. It is estimated that over 100 fungi will be discovered that are new to science. Recognition of these species will greatly inform broader research areas. Key features of the project include student training, internet-based information sharing, and public outreach.
Cryptic specialization in ectomycorrhizal fungi: Porcini mushrooms as a case study
with Laura Martínez-Suz (Kew), Bill Amos (Cambridge Univ.), Alex Papadopolus (Kew), Nathan Smith (Kew/Cambridge Univ.)
Boletus edulis is collectively a host generalist, but is that just the sum of a mosaic of cryptic specialists?
Resource partitioning is both cause and consequence of the evolution of increasingly specialized populations and a principal process that generates biodiversity (Thompson, 1994; Futuyma, 1998). Specialists are conventionally thought to evolve from generalists (Futuyma and Moreno, 1988), although exceptions to this prediction are well-documented (Futuyma and McCafferty, 1990; Lanyon, 1992; Thompson, 1994). Theoretically, extreme specialists require the evolution of highly specific adaptations, resulting in canalization that can make shifts to new resources increasingly unlikely. For this reason, specialists are sometimes believed to be both evolutionary dead-ends and more prone to extinction, but there is no general consensus over these hypotheses (Mayr, 1942; Futuyma and Moreno, 1988; Thompson, 1994).
The mycorrhizal mutualism, an association between the roots of plants (“hosts”) and fungi, in which the fungi exchange minerals extracted from the environment for fixed carbon from the host, is one of the most abundant and widespread examples of coevolution. Host-specificity of mycorrhizal fungi has long been debated, but the prevailing view is that the coevolutionary processes and patterns of mycorrhizal associations are diffuse (Futuyma, 2005), resulting in poor co-phylogenetic prediction of the symbionts and host, supporting the view that most mycorrhizal fungi are generalists.
Boletus edulis is a well-known and widely distributed obligately mycorrhizal mushroom that associates with a broad diversity of plant hosts. Superficially this appears to be a classic example of a highly successful generalist symbiont. However, host associations are mediated by interactions between individuals, yet no data exist on phenotypic or genetic variation at the individual or population level for B. edulis. This project aims to analyze genetic variation correlated with different hosts using whole genome associations from wild and captive individuals of B. edulis across it’s native range.
Collections-based discovery of fungal diversity
This is an ongoing project that combines fieldwork around the world (especially in the tropics) with comparative molecular and morphological analysis to discover, document, and describe fungal diversity. Physical vouchers (specimens) are at the core of this work, ensuring long-term scientific value by enabling reproducibility and reciprocal illumination. DNA sequences are the primary source of data used for identification and phylogenetic inference to reconstruct evolutionary history.
Individualism in biology: is the ‘Humongous Fungus’ the largest individual in the world?
Armillaria puigarrii. Costa Rica.
The ‘Humongous Fungus’ (Armillaria ostoyae) is thought to be an individual organism covering an area in excess of 2,300 acres in the Malheur National Forest, Blue Mountains, of eastern Oregon, USA (Ferguson et al. 2003, Schmitt & Tatum 2008). Evidence used by Ferguson et al. (2003) to determine individuals from multiple samples of A. ostoyae was based on somatic incompatibility (SI) of pairings from 108 isolates. But somatically compatible isolates are not necessarily identical genetically. Thus, the possibility exists of somatic compatibility between individuals of different genotypes, muddying the water of individuality in the Humongous Fungus. Moreover, spontaneous mutation may impact genomic composition in different parts of the individual, which has been documented in other long-lived individuals of Armillaria (Anderson & Catona 2014). Without detailed estimates of the true genomic diversity and similarity among the isolates it is not possible to say with great certainty that the Humongous Fungus represents a single, genetically homogenous individual, or if there is greater variation within an individual or among a colony of fragmented clones. The accumulation of genetic variation over the life of an individual can have phenotypic effects resulting in temporally differential selection. It could also result in a mosaic of selection across an individual because, in A. ostoyae, meiosis occurs in different areas of the fungus body where somatic mutations could be represented differentially. Genomic sequence data are needed to test these hypotheses, which is now possible using high-throughput sequencing technology that was not available in 2003. Unfortunately, many of the original isolates used in the 2003 study no longer exist (Ferguson, pers. comm.), requiring that new samples be acquired. This project proposes to recollect isolates of the Humongous Fungus to provide material that will be used to generate a high quality de novo genome assembly and resequencing from multiple isolates to observe individual genomic variation within and among somatic and reproductive tissues.
The diversity and evolution of coral mushroom crops in the fungus-farming ant mutualism
Pterula verticillata. Sarawak.
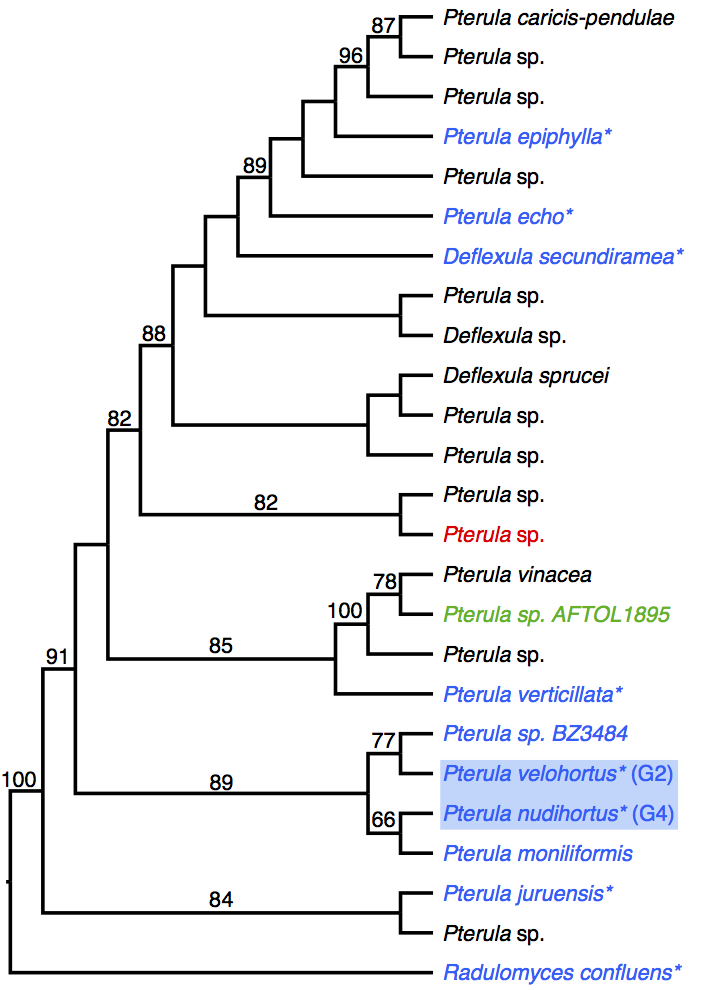
Approximately 20 million years ago an ancestor of the fungus-farming ant Apterostigma “pilosum group” (sensu Lattke 1997) adopted a novel fungal cultivar that now forms two phylogenetic clades of asexual fungi (“Group 2” and “Group 4”), which are only distantly related to the G1 and G3 Leucoagaricus cultivars (Chapela et al. 1994; Mueller et al. 1998). The G2 and G4 fungi were allied with the agaric family Tricholomataceae (Chapela et al. 1994; Mueller et al. 1998; Villesen et al. 2004), until recent phylogenetic evidence indicated that instead they share a recent common ancestor with the thread-like coral mushroom family Pterulaceae (Munkacsi et al. 2004, Schultz and Brady 2008, Dentinger et al. 2009). These two cultivated lineages were recently given formal scientific names, Pterula velohorta and P. nudihorta, respectively (Dentinger 2014), although they may be composed of more than one cryptic taxon.
The Pterulaceae are largely tropical, with only a few species reaching the northern and southern temperate latitudes. The Neotropics harbor the most species (42), 69% of which are endemic. The Afrotropics and Tropical Australasia are comparatively poorly sampled. The family was first formally recognized by Corner (1950), who admitted 6 genera (Corner, 1970) after Donk (1964) : Actiniceps Berk. & Broome (=Dimorphocystis Corner; 8 spp.), Allantula Corner (1 sp.), Deflexula Corner (16 spp.), Pterula Fr. (58 spp.), Pterulicium Corner (1 sp.), and Parapterulicium Corner (3 spp.). Recently, Dentinger and McLaughlin (2006) examined the phylogenetic distribution of club and coral (“clavarioid”) fungi with molecular phylogenetic methods and showed that Actiniceps is distantly related to other Pterulaceae. The genera Allantula and Parapterulicium have been unrecognized until very recently (C. Leal-Dutra et al., unpubl.).
Ongoing research on the diversity and molecular phylogenetics of Pterulaceae, particularly in the Neotropics, will shed light on the evolutionary history and origin of the fungus-farming ant cultivars. We are also using whole genome sequencing to build a phylogenomic tree and for comparative genomics that test hypotheses about the underlying genetic basis of domestication.
Are coral fungus symbionts domesticated crops?
Comparative genomic analysis of Pterula cultivars and 4 free-living relatives does not indicate a preponderance of derived gene families, or a large set of uniquely shared gene families, in the putative domesticates.
Domestication of plant crops provides one of the greatest illustrations of the power of selective forces in shaping organisms, spurring Darwin to use domesticated examples exhaustively in his argument for evolution and later in his thorough interrogation of the nature of genetic variation. Humans adopted agriculture around 10,000 years ago through domestication of wild plants followed by rapid selection for an assortment of desirable traits. Like human agriculture, the fungus-farming ant mutualism is thought to involve domestication of wild crops that have the potential to undergo ‘artificial’ selection3. Thus, a number of parallels have been drawn between human agriculture and ant fungiculture, but a distinguishing feature of the ant system is the complex crop-microbe community that appears to function synergistically to create a sustainable practice of crop monoculture. This emerging realization, along with the 50 million year history of successful ant fungiculture, has direct implications for improving human agricultural practices, from biological pest control to enhanced productivity from associated microbial diversity.



























 The Pterulaceae are largely tropical, with only a few species reaching the northern and southern temperate latitudes. The Neotropics harbor the most species (42), 69% of which are endemic. The Afrotropics and Tropical Australasia are comparatively poorly sampled. The family was first formally recognized by Corner (1950), who admitted 6 genera (Corner, 1970) after Donk (1964) : Actiniceps Berk. & Broome (=Dimorphocystis Corner; 8 spp.), Allantula Corner (1 sp.), Deflexula Corner (16 spp.), Pterula Fr. (58 spp.), Pterulicium Corner (1 sp.), and Parapterulicium Corner (3 spp.). Recently, Dentinger and McLaughlin (2006) examined the phylogenetic distribution of club and coral (“clavarioid”) fungi with molecular phylogenetic methods and showed that Actiniceps is distantly related to other Pterulaceae. The genera Allantula and Parapterulicium have been unrecognized until very recently (C. Leal-Dutra et al., unpubl.).
The Pterulaceae are largely tropical, with only a few species reaching the northern and southern temperate latitudes. The Neotropics harbor the most species (42), 69% of which are endemic. The Afrotropics and Tropical Australasia are comparatively poorly sampled. The family was first formally recognized by Corner (1950), who admitted 6 genera (Corner, 1970) after Donk (1964) : Actiniceps Berk. & Broome (=Dimorphocystis Corner; 8 spp.), Allantula Corner (1 sp.), Deflexula Corner (16 spp.), Pterula Fr. (58 spp.), Pterulicium Corner (1 sp.), and Parapterulicium Corner (3 spp.). Recently, Dentinger and McLaughlin (2006) examined the phylogenetic distribution of club and coral (“clavarioid”) fungi with molecular phylogenetic methods and showed that Actiniceps is distantly related to other Pterulaceae. The genera Allantula and Parapterulicium have been unrecognized until very recently (C. Leal-Dutra et al., unpubl.).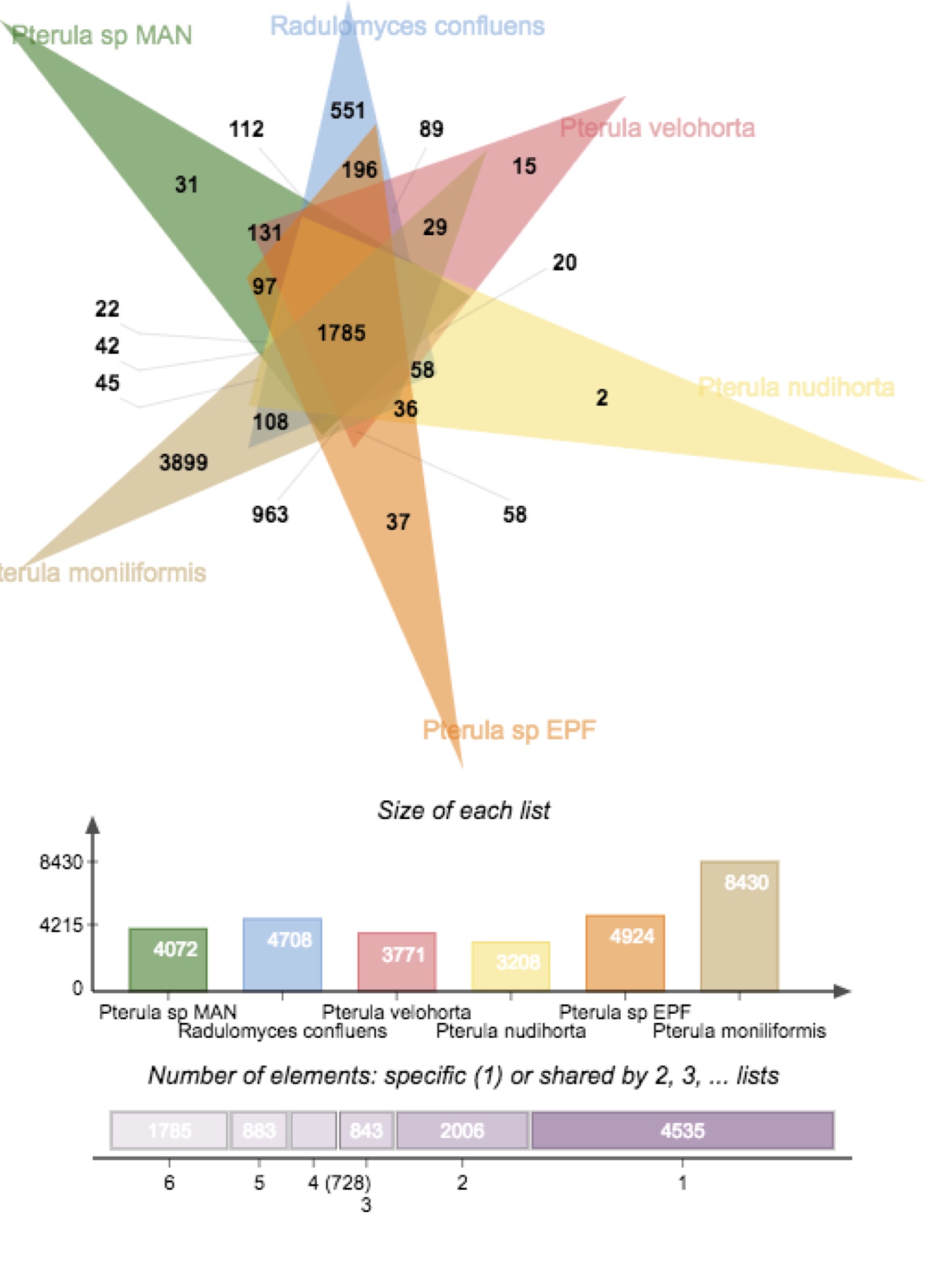


 Coming soon…
Coming soon…
